Age Determination
Two approaches are commonly used to determine an animal’s age at death. One deals with long bone fusion while the other focuses on dentition. Both have their pros and cons, and it is advised that data from both be presented whenever possible (Crabtree 1990).
Long bone fusion rates published by Silver (1969) serve as the standard reference for animal bone archaeologists. Fusion rates of the epiphysis at the end of a bone, or epiphyseal fusion, vary by bone, location of fusion area (top or bottom of bone), and species. Epiphyseal fusion is a widely used method among practicing zooarchaeologists, but it is not without its problems. Fusion rates are relatively imprecise and the affects of varying nutrition are poorly understood (Davis 1987:39). Crabtree (1990:162) also notes that in addition to nutrition, different breeds and castration also influence fusion rates. Fusion data have limited applicability in that they cannot assess the age of an animal after the latest point of fusion. For example, the latest fusion rates from sheep occur on the proximal humerus at 3.5 years (Silver 1969:252); therefore, once all centers have already fully fused, this method cannot be used to calculate advanced ages.
Ovicaprine fusion rates are calculated by grouping those bones that fuse at approximately the same stage in the animal’s life. The number of fused bones is divided into the sum of the fused and unfused bones for a particular age class, and is then expressed as a percentage. This value represents the frequency of ovicaprines surviving beyond a particular age class. An example drawn from the raw scores for the ovicaprine fusion data is illustrated in Table 2. For example, 22 bones are associated with centers that fuse at around 10 months old. The value is then divided into the total number of bones (34) for this particular age class. The result shows that 64.7% of the ovicaprines survived their first 6-10 months of life. The sum of the percentage values does not equal 100 because each age class is treated as a self-contained data set and is considered individually from other age categories. Therefore, the percentage values are specific to the age group in which they appear.
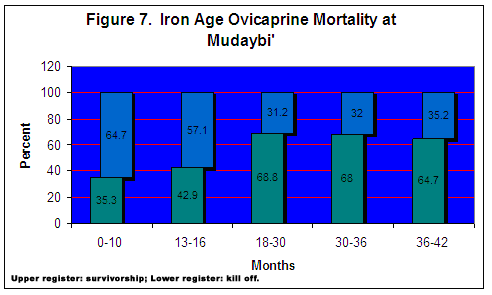
A total of 122 proximal/distal bone ends were used to compute ovicaprine mortality from the 8th century Moabite Fortress at Mudaybic, presented in Figure 7 . Each bar is divided into an upper and lower register; the upper represents the remains of those animals surviving beyond that particular age class, while the lower value represents the remains of animals killed within that age category. The observed mortality profile is considered in light of expected profiles that are product specific, that is, whether animals were raised for meat or dairy products. Meat producing economies with sheep normally kill young males once they achieve optimal weight gain, after which time the added bulk they accumulate is disproportionate to the amount of fodder required. Slaughter, then, is an economic consideration as it seeks to establish balance between the greatest bulk of the animal and the amount of food needed to achieve it. Animals are usually killed in their second or third year of life, while only a few males surviving into their later years are used for breeding (Payne 1973:281). As seen in the kill off rate for age classes corresponding to animals in their second and third year peaks, albeit only marginally, at 18-30 months. Many animals are also culled from the herd from earlier (10 months) and later (36-42 months). Significant kill off peaks at different times could indicate interest in both meat and dairy production, and perhaps some of the remains from older animals represent a few milk producing females. Since the body part distribution (below) indicates whole animals were not regularly kept at the site, exploitation strategies probably did not focus on large scale dairy production.
Dental attrition rates were defined by Payne (1973) and Grant (1982). Codes for some of the main domesticated animals, such as cattle, sheep, goats, and pigs, have been determined to correspond to a specific age or stage. The state of dental wear is assumed to reflect the age of the animal; teeth from the oldest individuals will exhibit the greatest amount of attrition, whereas those from younger individuals exhibit less wear and are more often characterized by cusps in pristine condition.
Payne’s model (1973) can be used to score the teeth of sheep and goats. His method specifically applies to mandibular molars and premolars. The method also requires that mandibular teeth be still set into the jaw rather than recording wear stages for isolated lower teeth. The reason is that scores for each tooth are considered as a series of datum points. An animal’s age at death can be more accurately estimated if the attrition scores from two or more teeth can be recorded from the same jaw, converging on a relatively small age range. Using isolated teeth results in less accurate estimates for age at death, expressed as greater age ranges, thereby inhibiting interpretation of the data. In Figure 8 we see two halves of ovicaprine mandibles from two different animals. Based on the wear on the chewing surface of the teeth, which mandible is from the older animal? Check for the answer at the end the article following the works cited.
It should be noted that not all kinds of tooth wear are directly linked to an animal’s age. Grant (1978:104) noted abrasive foods can influence the rate and nature of wear, as can soil types with more grit and greater acidity. Also, if an animal experienced injury or an abscessed tooth, it may prefer to chew on the unaffected side. By avoiding irritation, this would artificially inflate the degree of wear on the opposite overused side of the jaw. Premature incisor loss will also affect an animal’s ability to graze, thereby influencing tooth wear.
Dental eruption sequences are also used for determining age and are species specific (Silver 1969). This method is especially useful for recognizing ages for juveniles and sub-adults. However, it should be noted that poor nutrition can affect an animal’s tooth eruption. One of the limitations of dental eruption sequences is that it cannot be applied to species with rootless incisors (Hesse and Wapnish 1985:76). Incisors without closed roots continue to grow throughout the life of the animal, as enamel is constantly accreted to the base of the crown (Davis 1987:42).
There are advantages to relying on dental information for establishing an animal’s age at death, which have been clearly outlined by Payne (1973:281, 1985:145) and Klein and Cruz-Uribe (1984:43). Dental information can identify the presence of the young and the very old in a herd, since teeth continuously record the age of an individual throughout the course of its life. Due to this biological fact and that teeth are generally more durable and numerous than long bones, they are an excellent source of aging data. However, it is important to note that tooth wear tends to under-represent very young individuals (less than 6 months old); their teeth are not as durable and are more easily destroyed (Wapnish and Hesse 1988:89).